CONTRIBUCION DE LAS HOJAS AL LLENADO DEL RACIMO EN PLATANO (Musa AAB cv. Dominico-Hartón-Enano). Manuel Aristzabal L. MSc. y Rubén D. Landinez I.A.
INTRODUCCION.
La tasa de emisión de hojas, un factor clave en el desarrollo foliar del plátano, disminuye conforme la planta avanza en su crecimiento (Añstizábal et al., 1988) y depende de cada cutivañedad (Stover, 1979), de la temperatura ~Tumer, 1971) y del brillo solar (Hord y Speel, 1961). Dado que el plátano debe alcanzar cierto grado de desarrollo vegetativo para poder florecer y que dicha relación está dominada por el área foliar funcional (Champion, 1968), es de esperarse que el desarrollo foliar de la planta tenga gran influencia en la fructificación. Lambert (1976), considera que la diferenciación floral ocurre cuando la planta ha emitido un número ato de hojas funcionales. Quizás estos hechos sustentan la relación inversa entre el número de hojas emtidas a floración y el peso del racimo, observada por Martínez (1984), en plátano Dominico-Hartón y por Aristizábal et al. (1992), en el clon Dominico-Hartón-Enano-
Como la emisión foliar culmina con la floración, es de suponer que el número de hojas que la planta posea al momento de la floración es determinante para el llenado del racimo. A este respecto Belalcázar et al. (1991), establecieron que si la planta conserva 16 hojas durante todo el ciclo, puede perder hasta un 50% de las mismas sin que se afecte el peso del racimo. Martínez (1984), por su parte, había establecido que la planta requiere un mínimo de ocho hojas en floración para formar un racimo de buena calidad.
Del total de hojas que la planta presenta al momento de la floración, unas emergen del cormo y otras del tallo floral ; no obstante, aún no se ha establecido cuál es la contribución de unas y otras al llenado del racimo, lo que constituye el objeto de este estudio.
MATERIALES Y METODOS.
El experimento se realizó en la granja Montelindo (Región Santágueda) ubicada a 1.050 m-s-n-m- Durante el pido del cutivo la temperatura promedio fue de 23.6°C, la humedad relativa del 76% y la preciptación de 1.903 mm. Se sembraron 126 agujasrecoñadas del clon Dominico- Hartón-Enano a 3 m entre surcos y 2 m entre plantas.
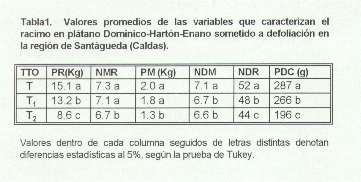
Dado que observaciones previas realizadas por Añstizábal permitieron establecer que del tallo floral del clon Dominico-Hartón-Enano emergen seis hojas (que en adelante se denominarán "Internas"), se efectuaron los siguientes tratamientos a tres grupos de 42 plantas cada uno. Testigo: Se dejaron todas las hojas que la -planta presentaba en floración y que en promedio fue de 12 hojas. Este grupo se empleó como Control m. Tratamiento 1 : Se dejaron las seis hojas "Internas" y se eliminaron las restantes, que se denominaron "Externas" ~Ti). Tratamiento 2 : Se dejaron las hojas "Externas" y las dos últimas ( Bandera y Penútima) de las hojas "lntemas", eliminando las cuatro restantes ~T2). Los tratamientos fueron realizados al momento de la emisión de la inflorescencia (Bellota).
Durante la cosecha se registraron el número de manos por racimo (NMR), el peso de este (PR), el número de dedos por racimo (NDR) y por mano (NDM), el peso de cada mano (PM) y el peso del dedo central de cada mano (PDC). Los valores obtenidos se sometieron a un análisis de varianza según un modelo completamente al azar con 14 replicaciones ~Tres plantas por replicación), antes de lo cual las variables NMR, NDR y NDM fueron transformadas a raíz cuadrada de cada valor. Las significancia estadística entre las diferencias de medias fue establecida mediante la prueba de Tukey a un nivel- del 5%.
RESULTADOS.
El análisis de varianza mostró diferencias significativas entre tratamientos para NMR y NDM y altamente significativas para las otras variables.
Las plantas testigo m presentaron los valores más altos de las variables que caracterizan el racimo y las del tratamiento 2 (T2) los más bajos. El NMR y el PM no difirieron estadísticamente entre T y Ti, mientras que el NDM fue estadísticamente igual entre Ti y T2. Los parámetros más afectados por los tratamientos fueron el PDC, el PM y el PR mientras que los menos afectados fueron el NMR, el NDM y el NDR.
DISCUSION.
Los resultados indican que la defolipción al momento de la floración puede reducir el peso y las características del racimo, y que la magnitud de dicha reducción dependerá del número y tipo de hojas eliminadas. Cuando se eliminaron de 6 a 7 hojas "Extemas" (Que se originan en la base del pseudotallo) y se dejaron las seis "lntemas" el peso del racimo disminuyó en un 12.6%; pero cuando se eliminaron cuatro de las seis hojas "Internas" (Que se originan a lo largo del tallo floral) y se dejaron las hojas "Externas", la penútima y la hoja bandera, la disminución en el peso del racimo fue del 43%. Esto permte suponer que el tipo de hoja eliminada y no la intensidad de la defoliación -como fue estudiado por Martínez (1984) y Belalcázar et al. (1991)- es lo que determina el efecto de la defoliación sobre el peso del racimo.
Como la intercepción de luz esta determinada por el tamaño, forma, posición y distribución de las hojas en la planta (Leopold y Kriedemann, 1975), es de esperarse que no todas las hojas contribuyan igualmente al llenado del racimo. De la información contenida en la Tabla 1 se deduce que las seis hojas que se originan en el tallo floral del plátano Dominico-Hartón-Enano aportan el 87.4% del peso total del racimo y que de estas las hojas bandera y penúltima contribuyen con el 44.4% y las hojas tercera, cuarta, quinta y sexta con el 43% restante. De hecho, el aporte de las hojas externas al llenado del racimo sería del 12.6%. Todo esto es válido si el racimo no aporta fotosintéticamente a su propio peso y si no ocurre traslocación de fotosintatos previamente acumulados en otras panes de la planta, aspectos estos que no fueron considerados en el presente estudio.
La alta contribución de las hojas bandera y penúltima al llenado del racimo se debe a que su posición en la parte más alta de la planta, les permite recibir más directamente la radiación solar, lo cual debe traducirse en mayor actividad fotosintética (Leopold y Kriedemann, 1975). También la posición relativa de cada hoja (fuente) con respecto al racimo (receptáculo) es otro factor que contribuye a explicar las diferentes contribuciones de las hojas al llenado del racimo, pues como ha sido demostrado en otras especies de plantas como el trigo (Rawson y Hofstra, 1968), las hojas que están más cerca a cada receptáculo (Espiga, mazorca y vaina, según el caso) aportan la mayor proporción de sus fotosintatos a su respectivo receptáculo. Esto explica el hecho de que las hojas bandera y penúltima, cuyo punto de inserción está más próximo al racimo, sean los que aportan la mayor contribución al llenado del mismo.
BIBLIOGRAFIA.
ARISTIZABAL, L-M- ; RODRÍGUEZ, L-R- y TABORDA, J-G- Análisis del crecimiento y la producción del plátano (Musa AAB C-V- Dominico) en función del clima. Manizales, Universidad de Caldas, 1988. Sin publicar
ARISTIZABAL, L-M- ; ARANGO, L-G- y CORREA, C-J- Relación del clima con el crecimiento y la producción del plátano (Musa AAB CV. Dominico- hartón Enano) en la región de Romelia Chinchiná (Caldas), Universidad de Caldas, Facultad de Agronomía, Mandales 1992. Sin publicar.
BELALCAZAR, C-S- , MERCHAN, M-V-, BAENA, A-H- y VALENCIA, MJ A. Efecto de la época y el grado de defoliación sobre la producción. Seminario de Actualización sobre la Investigación en el Cultivo del Plátano Ltografla Cafetera, Manizales, 1991 p. 54-59.
CHAMPION, J. El plátano. En : Agricultura tropical. Barcelona Blume, 1968 p. 48-70.
HORD, H. and SPEEL, D-P- Temperature as basis for forecasting banana production. In : Tropical Agriculture. Val. 39. (1961). p. 219 - 223.
LAMBERT, C. Caracteñstiques de croissance et les phases de developpement ches le bananier plantain (Var carne) signification physiolojique des caracteñstiques de croissance des sevilles fontiÓn elles Fruts (Francia) Val. 31, No. ll (1976) ; p. 678 - 683.
LEOPOLD, A-C- and KRIEDEMANN, P-E- Plant growth and developpment. New York : Mc Graw Hill Book CO., 1975.
MARTINEZ, GA. Determinación del área mInima foliar en plátano en el trópico húmedo. Revista ICA. Val. 19, No. 2 (1984) ; p. 183-187
RAWSON, H. M. and HOFSTRA, G. Translocation and remobillization of14-C assimilated at diferent stages by each leaf of the wheat plant. Australian Journal of Biological Science Val.12 (1968) p.371- 376.
STOVER, RH Pseudostem growth, leaf production and flower initiation in the Grand Nain banana. In : Tropical agncuture research services. Bulletin N>o.6. 1g7g. p.37.
TURNER, D-W- Eftects of dímate on rate of banana leaf production Tropical Agñcuture Val. 48, No. 3 (1971 ). p. 283 - 287.
![]()